[See
ref 15
]
What could be the biological function of cellular infrared 'vision'?
As shown in the flow diagram below, our work up to this point seems to link
cellular infrared 'vision' with animal development which depends critically on cell migration .

(For more explanations click the blue banners)
Therefore, we suspect that the infrared 'vision' of cells plays a role in
development.
There is an additional reason for this conjecture. Animal cell migration is very slow
and animal development occurs simultaneously everywhere in the embryo. What, if
a migrating cell makes a mistake and ends up in the wrong place? It cannot stop the
development everywhere else, turn back the developmental clock and start from scratch. Therefore,
it seems good engineering to provide the migrating cells with the means to 'look ahead' towards their
spatial goals.
How far should a cell look ahead? In a developing embryo any spatial goal is a moving target. If it takes a cell 6-10 hours to reach this goal,
it would probably no longer exist because development may have changed the area around the goal too much.
Since an animal cell needs about 1.5 hours to cross its own diameter of appr. 20 µm its spatial goals
should be no further away than 4-6 cell diameters or 80-120 µm. Indeed, our experiments showed that
the cells ignored microscopic light sources that were further away than about 60 µm [
ref 13.
].
Cells detect each other at a distance.
It stands to reason that the major kinds of spatial goals of cells in an embryo are other cells.
For many years I oberved that approaching cells reached out towards each other from a distance,
as if they 'saw' each other.
The figure below shows a remarkable example.
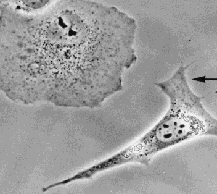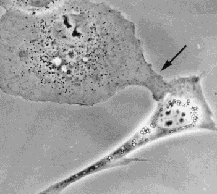
(The illustration is animated.Click here for a minimal strip of frames.)
In this case an epithelial cell from an African Green Monkey (upper left corner)
extends in a very targeted way towards a fibroblast from a mouse that had turned its front
towards the epithelial cell at an earlier time. In spite of the differences of animal and tissue type, the 2 cells
recognized their presence at a distance and actively approached each other.
Cells react to each other across a film of glass.
In order to test whether such cell-to-cell encounters were mediated by some chemicals, I
separated cells by a thin, but chemically impenetrable glass film.
The figure below shows a Hamster fibroblast sitting on one side of the glass film (Let us call it the 'A'-face).

On the other side of the glass film (Let us call it the 'B'-face) we forced cells to grow in
a pattern of parallel stripes (see green lines on the figure below).

(The illustration is animated.Click here for a minimal strip of frames.)
We found that the cells on the 'A'-face oriented themselves predominantly perpendicular to the
cell stripes on the 'B'-face (see red lines on the above figure), even though the 2 faces were insulated from each other by the
impenetrable and inert glass film. When we coated the 'A'-face with an opaque gold film and then
tried the experiment, no such special orientation was found. On the other hand, if we coated it with a film
of silicone which is opaque for visible light and transparent for near infrared light, the special
orientation of the cells reappeared. In other words, only as long as infrared light could
pass through the glass film, the cells were also able to determine the direction of the cell stripes on the
opposite face.
Significance for cell intelligence:
Infrared 'vision' may represent a novel form of cell-to-cell communication
The observation of a perpendicular orientation between cells in different planes is not
the amazing results. Many tissues in the body (e.g. the cornea, the intestines and many others)
are weaved like this. Cells seem to be
programmed to line up side-by-side in the same plane and perpendicular to each other in the planes
above and below. The amazing result is the observation that the cells on the 'A'-face knew the orientation of the
cells on the 'B'-face if near-infrared light could pass across the glass film. Similar to the
argument about cells following guiding 'roads'
the cells on the 'A'-face had to 'see' at least 2 different points on the cell stripes on the
other side of the glass film in order to determine the direction of the stripes. In the above experiment, however,
we did not provide any pulsating infrared light. Therefore, if the cells themselves were the sources of the light
that passed through the glass, they were communicating their spatial orientation to the cells in a
layer above. Therefore, it seems possible that the infrared 'vision' of cells is used in the developing
animal to weave tissues.